Why we don't yet have evidence for macrocolumn-scale structure/function throughout all of Cortex?
The implicit belief of neuroscience over the past 100 years has been the neuron doctrine which says that the single neuron is the fundamental computational unit of the brain, or in particular, of the cortex. This has likely been conflated in minds with the idea that the single neuron is also the fundamental, or atomic, unit of representation, or meaning. Certainly, most neuroscientists historically have been thinking in terms of what individual neurons represent, often described as Gaussian or Gabor-like spatial functions (in any case, having a single mode) in one or a very few dimensions. Once you begin to think in terms of sparse distributed representations (SDR), a.k.a., cell assemblies, you can quickly come to understand that all of these low-dimensional Gabor-like receptive fields (RFs) that have been described for single neurons over the decades are simply artifacts of the experimental (mostly, single/few-unit electrophysiological) methodology. This view is supported, for example, by the results of Nandy et al (2013), which used much finer scaled stimuli. The essential properties of the traditional electrophysiological methodology have been:
- Recording one or few neurons, generally a mix of excitatory and inhibitory, within any given large volume at any one time
- Averaging over numerous trials (and thus, to various degrees, over different conditions), and
- Using simple (i.e., low statistical complexity), low-dimensional stimuli that are of larger spatial extent than the that of the single unit's RF.
As Rafael Yuste has said, this is like watching a movie through one pixel. I submit that once we have the ability to measure all cell activity (really only all the principal cells would be sufficient) in macrocolumnar-scale horizontal extents and layer-thickness-scale vertical extents, e.g., L2/3, and on the ms time scale, we will be able to see macrocolumner SDRs activating and deactivating, probably on the gamma time scale, perhaps also on the theta time scale. If we can determine the boundaries of a functional macrocolumn then this will be easy. However, it may well be that actual functional cortical macrocolumns can physically overlap; i.e., the same minicolumns, which we propose operate as winner-take-all (WTA) "competitive modules (CMs), may physically be part of multiple subsuming macrocolumns, only one of which will be active at any given moment (i.e., gamma cycle), as suggested in Figure 1. This, in conjunction with the experimental limitations mentioned above, are likely why we have not thus far observed functional macrocolumn-scale cell assemblies in much of cortex, especially more frontal regions. Hopefully, rapid advances in calcium imaging and voltage senstive dyes will redress this situation soon.
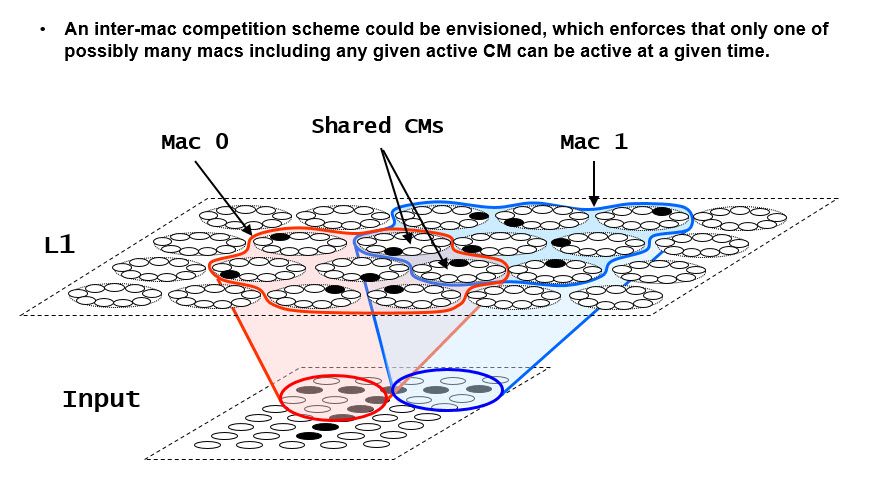
Figure 1: Illustration of the concept that real macrocolumns ("macs") could physically overlap, sharing minicolumns (i.e., CMs). Sparsey already assumes a WTA competition mechanism within a CM. There is nothing in the theory that would prevent an additional meta-level of competition ensuring that at any given time, only one of the perhaps multiple macs that include any particular CM can be active. In this case, only one of Mac 0 or Mac 1 could be active at any given time, e.g., in any given gamma cycle. The main point is that if we were (electrophysiologically) observing a given cell within one of the shared CMs across many different trials (possibly involving a wide range of stimuli), that cell might be activating sometimes as part of one subsuming mac and sometimes as part of another. This would make functional mac borders quite hard to discern. And note that this variability across trials is in addition to the idea that that cell might activate as part of (perhaps many) different cells assemblies within any sone of those subsuming macs. Furthemore, although the RFs of the two macs in this case are disjoint, i.e., the red and blue ellipses in the input level, this is likely not the case in the brain, nor is it required by our theory.
Figure 2 shows a more complex example in which a field of CMs has some pattern of overlapping macs imposed upon it. Note that we show only the coding field (L1) in each of this figure's eight panels, not the input field (as in Fig. 1). A different color outline and shading is used for each mac. The cell indicated by the red arrow is part of four containing macs (green, purple, blue, and red). Reading down the left column of figures, we show the subject cell active in codes in each of those four macs. Each possible active subsuming mac imposes different constraints on what other macs can be active. In the top row, the indicated cell is active as part of a cell assembly active in the green mac. Given the assumption of a meta-level, i.e., inter-mac-level competitive circuitry, this means that no other mac that intersects with the green mac can also be simultaneously active (e.g., on the same gamma cycle). A possible global activity pattern, meeting that inter-mac competition constraint, is shown here, with the gray and orange macs also active. To the right, we show the global activity pattern with only the active mac colors included to show that they do not intersect. Of course, if the gray mac is active, then the pink mac cannot be and if the orange mac is active then the teal mac cannot be. In the second row, we show the same subject cell (red arrow) active as part of a cell assembly in the violet mac. In this case, no other mac that intersects with the violet mac (green, brown, gray, and orange) can simultaneously be active. Again, at right, we show a clearer picture revealing the three non-intersecting co-active macs, red, violet, and teal. The other rows have siimilar interpretations.
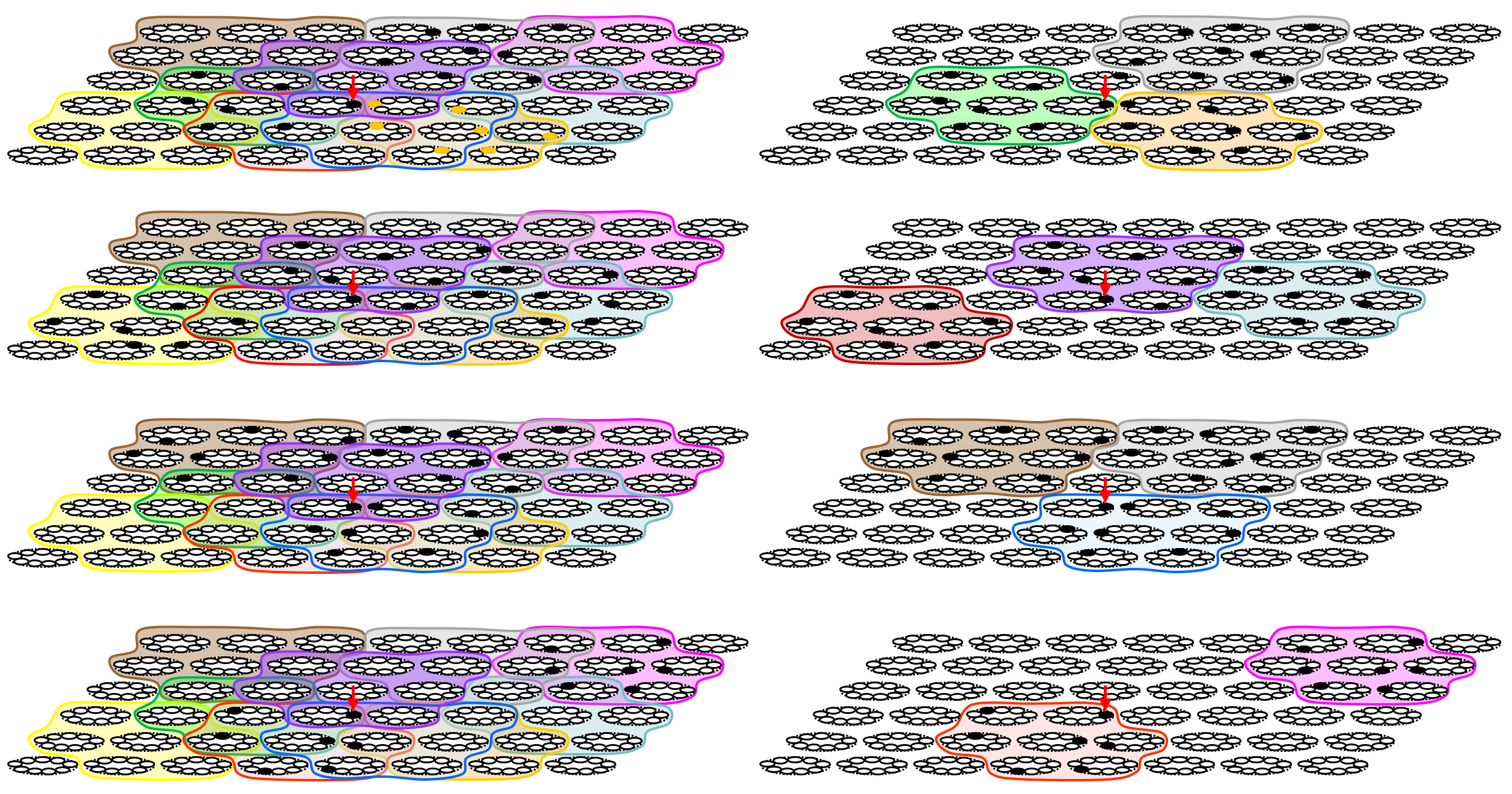
Figure 2: More complex example showing overlapping macs. The policy is that an active cell [and its subsuming WTA competitive module (CM), the proposed analog of the minicolumn] can be part of only one subsuming active mac at any one time. See text.
We include Figure 3 just to show what the situation looks like when there is no information about mac boundaries. The left column is the same as the right column of Figure 2. The right column is the same as the left but with the mac boundaries removed. We emphasize that if something like this overlapping mac scheme was operative in cortex, it would greatly complicate detection of functional macrocolumns. For example, if an electrophysiologist is measuring the red-arrow cell, he will typically be averaging over instances in which that cell is active in different subsuming macs. But the cell’s response, and its meaning, and indeed the portion of its overall RF (again, we are not showing the cells' RFs, i.e., as the input level, in these figures) that is actually controlling it in the given instance, will differ perhaps greatly, depending on which subsuming mac is active. We also emphasize that given our current quite limited understanding of the extremely complex inhibitory circuitry of cortex, such a scheme of multiple scales of superposed competition is well within the realm of plausibility.
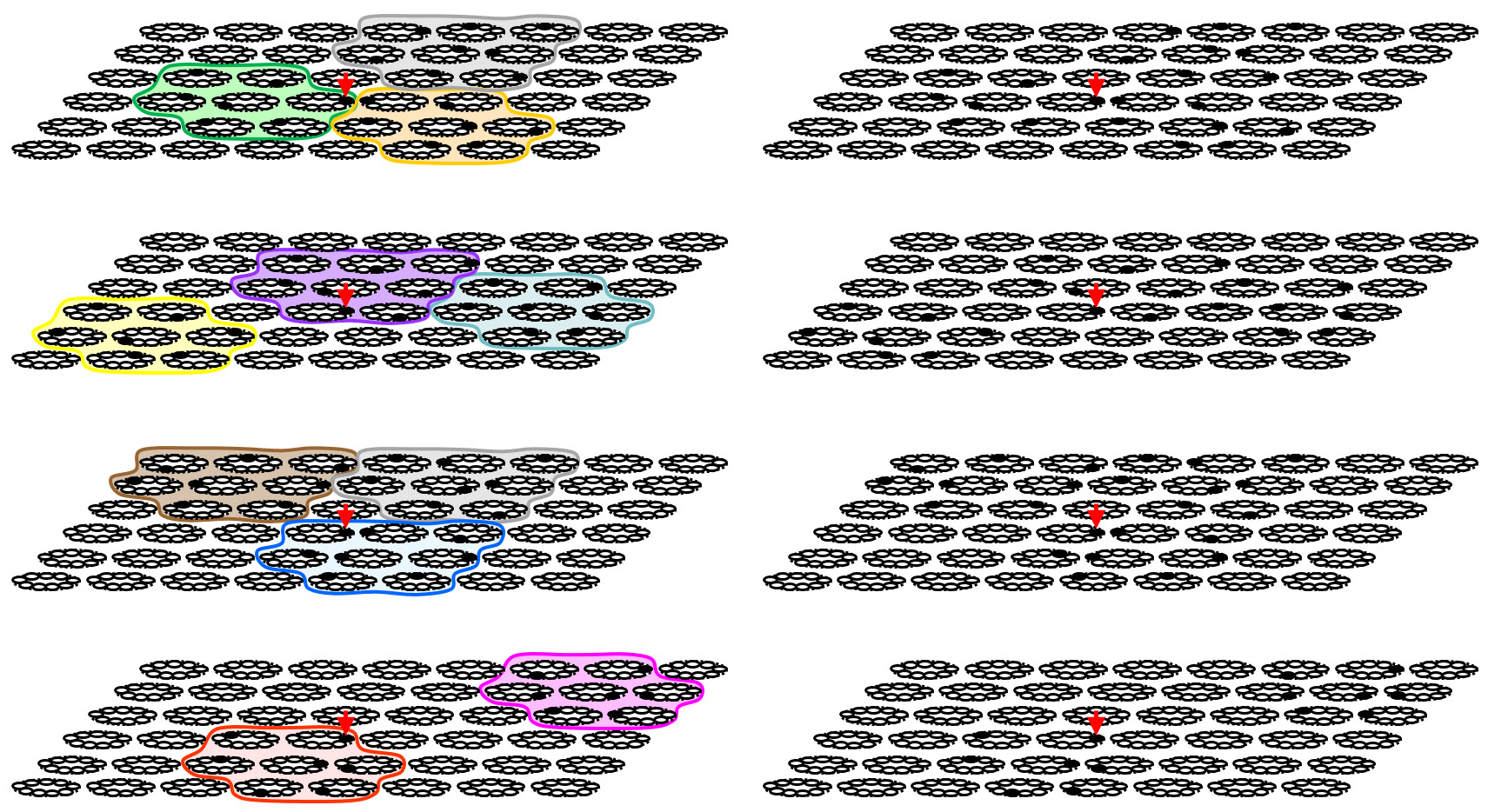
Figure 3: This figure is added just to show the situation when no information about subsuming mac boundaries is given (righthand column). This might correspond to something like what calcium imaging might show.